* フォトトロピンとは
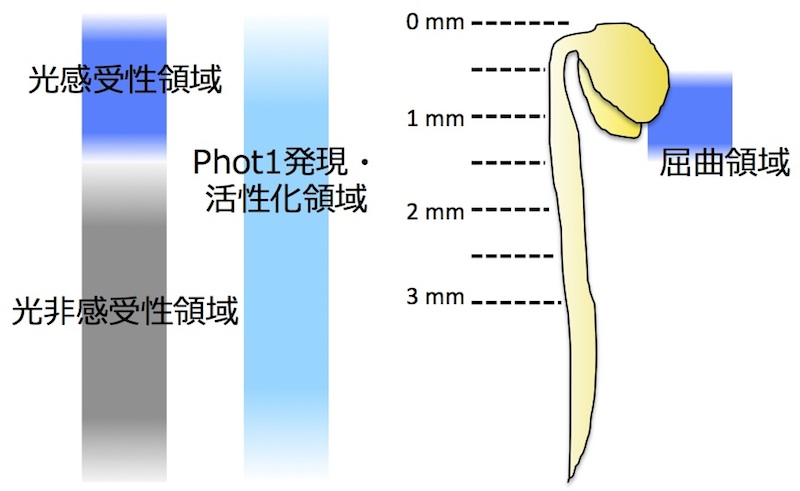
イネ科植物の幼葉鞘を用いて1880年にチャールズ・ダーウィンらが発見した「光屈性」を誘導する光受容体が「
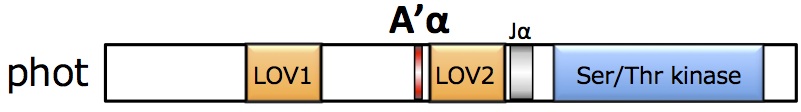
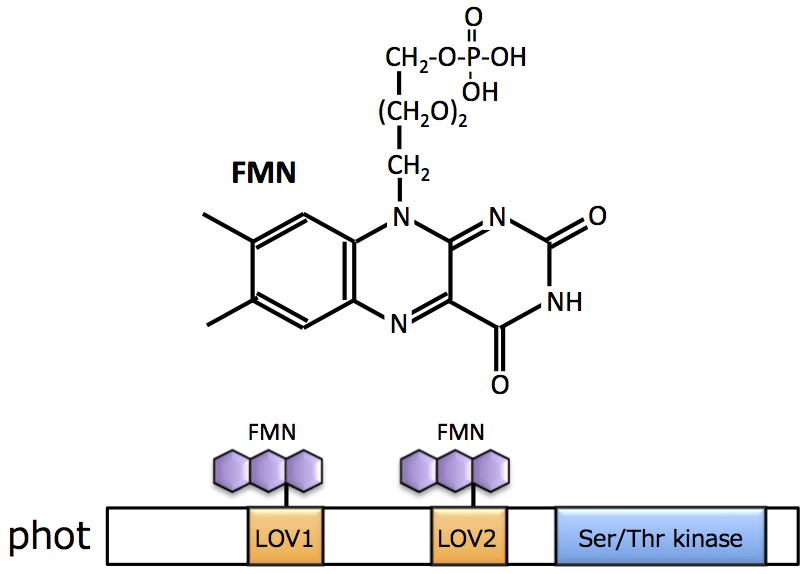
フォトトロピン(phot)」である。photはN末端側に光感知に関わるLOV(Light, Oxygen, Voltage)ドメインを2つ(LOV1、LOV2)と、C末端側にSer/Thrキナーゼドメインを有する。LOVドメインには発色団としてFMNが結合しており、青色光刺激によってLOVドメインを中心にphotタンパク質全体の構造変化が誘導されると、C末端側のキナーゼ活性が上昇し下流に情報が伝達される。 (
研究背景;光受容体 参照)。
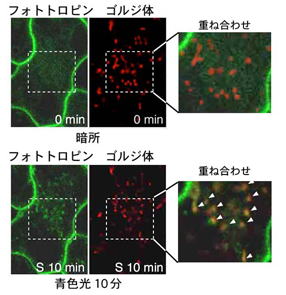
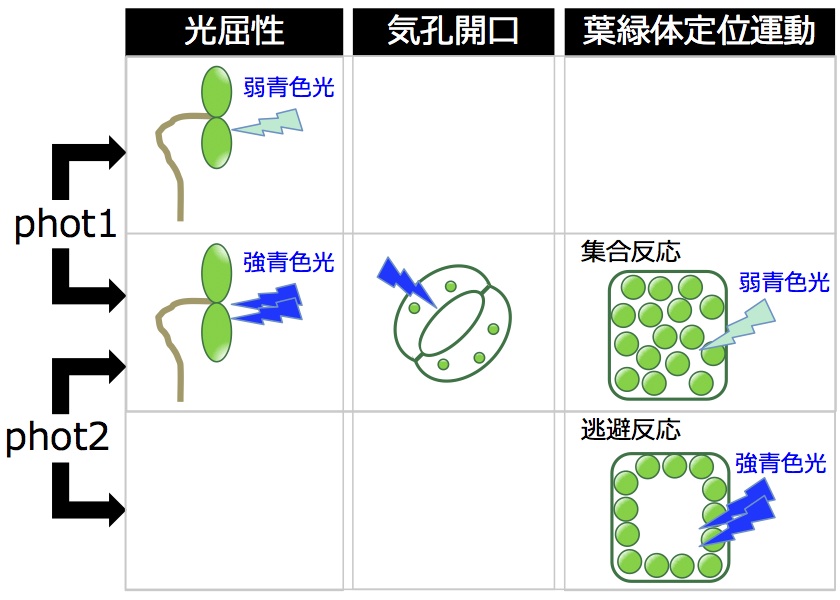
私たちが主に研究対象としている高等植物シロイヌナズナにおいて、フォトトロピンは光屈性以外に葉緑体定位運動(青色光に応じて細胞内の葉緑体の位置が変わる反応)、気孔開口、葉の扁平化、植物体の成長などの応答を制御する。シロイヌナズナには2つのphot分子種(phot1、phot2)が存在するが、この2つのphotは基本的には冗長的に働く。強青色光による光屈性、弱青色光に対して葉緑体が集まる反応(葉緑体集合反応)、気孔開口などに対する制御がそれである。一方、弱青色光での光屈性はphot1特異的反応であり、強青色光に対して葉緑体が逃げる反応(葉緑体逃避反応)はphot2特異的な反応である。
 * 研究の目的と内容
* 研究の目的と内容
フォトトロピン情報伝達においては、キナーゼ活性によるリン酸化反応が必須である。photのリン酸化基質としては既に幾つかの因子が既に同定されているものの、フォトトロピン情報伝達系の全容は未だ明らかとなっていない。
私たちの研究室では、生理学・生化学・細胞生物学・分子生物学・遺伝学的手法を駆使することで
「フォトトロピン分子の特性」や
「フォトトロピン情報伝達に関わる場と因子」を明らかにしたいと考えている。これまでに、分子生物学的手法を中心に、植物シロイヌナズナのみならずクラミドモナス(緑藻)・大腸菌・酵母・培養細胞など様々な生物でphot分子を発現させることで独自の解析手法を確立してきた。研究内容の詳細については下記項目を参照して頂きたい。
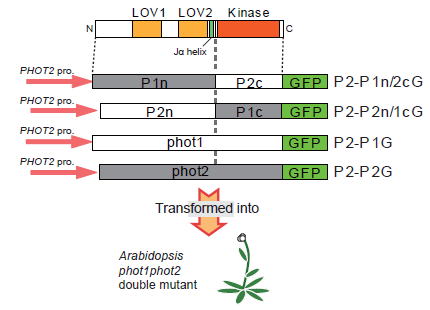
1.フォトトロピン分子の機能ドメイン
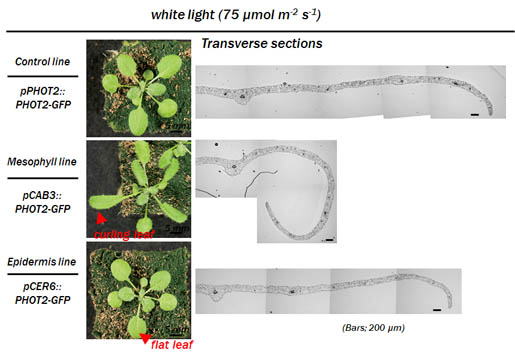
2.フォトトロピン応答の組織・器官間シグナル伝達
3.フォトトロピン情報伝達経路
4.フォトトロピンを利用したオプトジェネティクス
これら基礎研究を通して「
植物が獲得した巧妙なメカニズムを分子レベルで知る」ことが、農学・医学・工学などの様々な応用分野への礎となると期待する。オプトジェネティクス研究もその最初のステップである。