|
丂丂僼傿僩僋儘儉偺庡梫側暘巕庬偲偟偰phyA偲phyB偑抦傜傟傞丅変乆偼丄僎僲儉忣曬偑傑偩惍傢側偄帪戙偐傜偙偺栤戣偵庢傝慻傫偱偒偨丅傑偢丄僔儘僀僰僫僘僫傪嵽椏偵phyB曄堎懱偺摨掕偵婑梌偡傞偲偲傕偵丄phyA偺曄堎懱傪帺傜扨棧偟偨丅偝傜偵丄僩儅僩傗僄儞僪僂偺僼傿僩僋儘儉曄堎懱偺夝愅傪僆儔儞僟偺僌儖乕僾偲嫟摨偱峴偭偨乮van Tuinen et al., 1995a, b, 1996; Weller et al., 1995; Kerckhoffs et al., 1997側偳乯丅
丂丂丂丂丂丂丂丂丂丂丂丂 僼傿僩僋儘儉寚懝曄堎懱
丂丂丂 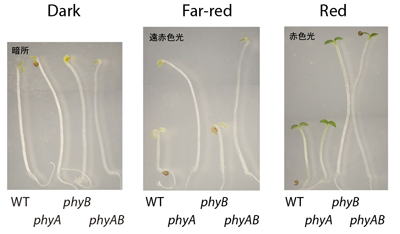
丂丂丂丂丂乮挿扟嶣塭乯
丂丂phyA偲phyB偺曄堎懱傪梡偄傞偙偲偵傛傝丄偦傟偧傟偺暘巕庬偺惗棟揑側栶妱傪柧傜偐偵偡傞偙偲偑壜擻偵側偭偨丅偙傟傪棙梡偟偰丄岝敪夎偵偍偄偰phyA偲phyB偺墳摎偺嶌梡僗儁僋僩儖偑戝偒偔堎側傝丄phyA偑屆偔傛傝抦傜傟傞挻庛岝斀墳偺岝庴梕懱偱偁傞偙偲傪徹柧偟偨乮Shinomura et al., 1996乯丅傑偨丄偙傟傜偺曄堎懱傪梡偄偰丄僼傿僩僋儘儉偺崻偺椢壔偵偍偗傞栶妱乮Usami et al., 2004; Usami et al., 2007乯丄梩偺滸暯壔偵偍偗傞栶妱乮Kozuka et al., 2013乯側偳傪帵偟偰偒偨丅
丂丂偙傟傜偺曄堎懱偼丄僼傿僩僋儘儉尋媶偵寚偐偣側偄帒尮偲峀偔擣幆偝傟傞偵帄偭偨丅変乆偼丄偙傟傜偺曄堎懱傪梡偄偰丄條乆側暘栰偺尋媶幰偲岝墳摎偵娭偡傞嫟摨尋媶傪峴偭偰偄傞乮Batschauer et al., 1996; Genoud et al., 1998; Cerdan et al. 1999, Yamamoto et al., 2003; Tamura et al., 2003; 懠懡悢乯丅
丂丂 僼傿僩僋儘儉 尋媶top傊
|
丂丂僼傿僩僋儘儉偼悈梟惈偱偁傞偙偲偐傜丄挿偄娫嵶朎幙偱婡擻偡傞偲峫偊傜傟偰偄偨丅偟偐偟変乆偼丄GUS梈崌僞儞僷僋幙傪梡偄偰丄phyB偺C-枛抂僪儊僀儞偵妀堏峴妶惈偑偁傞偙偲傪弶傔偰帵偟偨乮Sakamoto & Nagatani, 1996a乯丅偝傜偵丄GFP傪梡偄偰姰慡挿偺phyB偑岝偵墳摎偟偰妀撪偵堏峴偟丄妀撪偱梓棻忬偺峔憿懱傪宍惉偡傞偙偲傪帵偟偨乮Yamaguchi et al., 1999乯丅傑偨丄擔棫惢嶌強偺僌儖乕僾偺屆扟傜偺僌儖乕僾偲嫟摨偱丄僄儞僪僂偵偍偄偰phyA偑岝埶懚揑偵妀撪偵堏峴偡傞偙偲傪柶塽壔妛揑偵偟傔偟偨乮Hisada et al., 2000乯丅偙傟傜偺尋媶偵傛傝丄僼傿僩僋儘儉偺僔僌僫儖揱払偵偍偗傞廳梫側僗僥僢僾偑柧傜偐偲側偭偨丅
丂丂 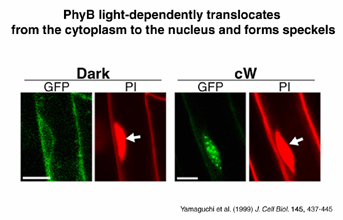
丂丂乮幨恀偼Matsushita et al., 2003偵傕偲偯偔乯
丂丂忋婰偺尋媶偑峴傢傟偨偺偲偪傚偆偳摨偠崰丄暷崙偺Quail傜偺僌儖乕僾偵傛傝丄僼傿僩僋儘儉偑揮幨場巕偲暔棟揑偵憡屳嶌梡偡傞偙偲偑帵偝傟丄僼傿僩僋儘儉偼妀撪偱僔僌僫儖傪揱払偡傞偲峫偊傜傟傞傛偆偵側偭偨丅偝傜偵嵟嬤偱偼丄phyA偑妀撪偱儐價僉僠儞壔偝傟傞偙偲傪帵偡嫟摨尋媶傪峴偄丄妀偑phyA偺暘夝偺応偱傕偁傞偙偲傪帵偟偨乮Seo et al., 2004乯丅傑偨丄PIF偺敪尒幰偱偁傞Quail傜偲嫟摨偱丄偲僼傿僩僋儘儉偲PIF偺憡屳嶌梡偵娭偡傞嫟摨尋媶傪峴偭偨乮Kikis et al.k 2009乯丅
丂丂 僼傿僩僋儘儉 尋媶top傊
|
丂丂僼傿僩僋儘儉偺N-枛抂懁僪儊僀儞偼敪怓抍傪寢崌偟岝偵墳摎偟偨Pr-Pfr宆岝曄姺傪帵偡丅堦曽丄C-枛抂懁僪儊僀儞偼俀検懱壔傪扴偆丅偝傜偵丄C-枛抂懁偵僪儊僀儞撪偵丄PAS僪儊僀儞傗僉僫乕僛僪儊僀儞偑擣傔傜傟傞偨傔丄僼傿僩僋儘儉偼C-枛抂懁僪儊僀儞傛傝僔僌僫儖傪敪怣偡傞偲峫偊傜傟偰偒偨丅偟偐偟側偑傜丄C-枛抂傪扨撈偱敪尰偝偣偨怉暔偱尠挊側昞尰宆偼擣傔傜傟側偐偭偨乮Sakamoto & Nagatani, 1996b乯丅
丂変乆偼丄忋婰偺寢壥偐傜俈擭屻丄偙偺栤戣偵娭偡傞廳梫側寢壥傪摼偨丅phyB偺N-枛抂懁僪儊僀儞傪俀検懱壔偟偰妀偵嬊嵼偝偣傞偲丄傎偲傫偳惓忢側岝庴梕懱偲偟偰婡擻偟偨乮Matsushita et al., 2003乯丅偙偺寢壥丄塱擭偺乽忢幆乿偵斀偟偰丄C-枛抂僪儊僀儞偼岝僔僌僫儖偺揱払偵昁偢偟傕昁梫側偄偙偲偑柧傜偐偲側偭偨丅
丂丂 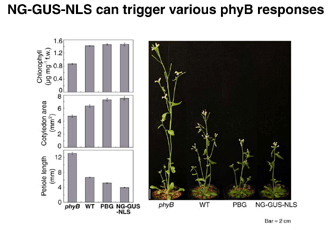
丂丂乮幨恀偼Matsushita et al., 2003偵傕偲偯偔乯
丂丂忋婰偺寢壥傪庴偗丄偝傜偵抁偔偟偨N-枛抂懁僪儊僀儞偺惗棟妶惈傪挷傋偨寢壥丄N-枛抂懁僪儊僀儞撪偺C-枛抂懁偵懚嵼偡傞栺200傾儈僲巆婎傛傝側傞PHY僪儊僀儞傪彍偄偰傕岝庴梕懱偲偟偰婡擻偡傞偙偲傪帵偟偨乮Oka et al., 2004乯丅懠偺尋媶僌儖乕僾偺寢壥偲崌傢偣傞偲丄妀撪偱偺僔僌僫儖揱払偵杮幙揑偵昁梫側偺偼丄僼傿僩僋儘儉傪峔惉偡傞栺1100傾儈僲巁巆婎偺偆偪傢偢偐350傾儈僲巁巆婎掱搙偲峫偊傜傟傞丅
丂丂丂 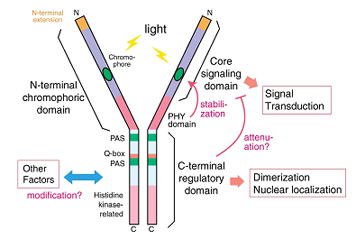
丂丂変乆偼丄N-枛抂懁僪儊僀儞撪偱phyB偺婡擻敪尰偵娭梌偡傞傾儈僲巁巆婎傪摿掕偡傞偨傔丄摉奩僪儊僀儞傪敪尰偡傞怉暔偵曄堎傪摫擖偟丄墳摎偑掅壓偟偨曄堎姅偺戝婯柾僗僋儕乕僯儞僌傪峴偭偨乮Oka, Matsushita et al., 2008)丅摼傜傟偨姅偺拞偐傜丄摫擖phyB抐曅偵傾儈僲巁抲姺偑擣傔傜傟傞傕偺傪慖暿偟丄偦傟偧傟偺曄堎phyB抐曅偺惈幙傪挷傋偨丅偦偺寢壥丄暘岝岝妛揑惈幙傪懝側偆曄堎偼丄N-枛抂懁僪儊僀儞撪偺僒僽僪儊僀儞偺偆偪丄GAF僪儊僀儞偲PHY僪儊僀儞偵廤拞偟偰偄偨丅堦曽丄僼傿僩僋儘儉偵摿挜揑側峔憿偱偁傞"light sensing knot"偺嬤朤偵丄暘岝岝妛揑惈幙傪懝側偆偙偲側偔僔僌僫儖揱払偵塭嬁偡傞暋悢偺曄堎傪尒弌偟偨丅偙偺寢壥偼丄僼傿僩僋儘儉偺N-枛抂懁僪儊僀儞偐傜偺僔僌僫儖敪怣偵"light sensing knot"偑娭傢傞偙偲傪帵嵈偡傞旕忢偵嫽枴怺偄寢壥偱偁傞丅
丂 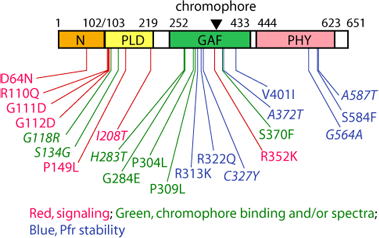
丂丂偝傜偵丄暷崙偺Quail尋偲嫟摨尋媶偵傛傝丄忋婰偺僔僌僫儖揱払偵娭傢傞傾儈僲巁巆婎偑丄僼傿僩僋儘儉偲憡屳嶌梡偟偰僔僌僫儖傪揱払偡傞bHLH宆偺揮幨場巕偱偁傞PIF3偲偺寢崌偵傕娭傢傞偙偲偑柧傜偐偲側偭偨乮Kikis, Oka et al., 2008乯丅廬偭偰丄僼傿僩僋儘儉偼N-枛抂撍弌晹丄PAS丄"light sensing knot"側偳偺摿掕偺昞柺峔憿傪夘偟偰僔僌僫儖揱払傪峴偭偰偄傞偲憐憸偝傟傞丅
丂丂 僼傿僩僋儘儉 尋媶top傊
|
丂丂変乆偼丄僼傿僩僋儘儉傪係偮偺晹埵偵傢偗丄phyA偲phyB偺娫偱偦傟偧傟偺晹埵傪岎姺偡傞phyA/phyB僉儊儔僞儞僷僋幙傪峔抸偟丄怉暔偵堚揱巕摫擖偟偰偦偺惈幙傪挷傋偰偒偨丅偦偺寢壥丄phyA偑帵偡條乆側摿挜偼丄堎側傞晹埵偺phyA攝楍偑寛傔偰偄傞偙偲偑柧傜偐偲側偭偨丅偡側傢偪丄phyA暘巕偼丄條乆側摿挜傪傕偮儌僕儏乕儖偑慻傒崌傢偝偭偨暘巕偲懆偊傜傟傞偙偲偑暘偐偭偨乮Oka, Ono et al., 2012乯丅
丂 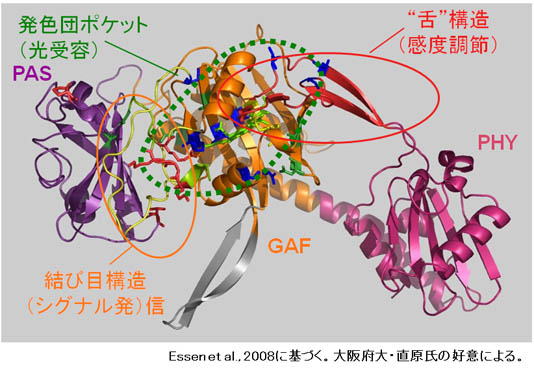
丂丂 僼傿僩僋儘儉 尋媶top傊
|
|
丂 僼傿僩僋儘儉偲憡屳嶌梡偟偰壴夎宍惉傪惂屼偡傞PHL
|
丂丂挿擔怉暔偱偁傞僔儘僀僰僫僘僫偵偍偄偰偼丄僼傿僩僋儘儉B乮phyB乯偵傛偭偰壴夎宍惉偑梷惂偝傟傞偙偲偑抦傜傟偰偄傞丅変乆偼丄嫗搒戝妛丒峳栘悞嫵庼丄墦摗媮彆嫵傜偲嫟摨偱丄偙偺惂屼偵娭傢傞怴婏場巕PHL傪摨掕偟丄偦偺婡擻傪徻偟偔挷傋偨丅偦偺寢壥丄PHL偑phyB偺婡擻傪惂尷偡傞偙偲偱壴夎宍惉傪憗傔偰偄傞偙偲偑暘偐偭偨丅傑偨丄phyB, PHL偵壛偊偰丄偦偺壓棳偱壴夎宍惉偺惂屼傪峴偭偰偄傞偲峫偊傜傟偰偒偨CO偺嶰庬偺僞儞僷僋幙偑妀撪偱暋崌懱傪宍惉偡傞偙偲傪帵偡寢壥傪摼偨乮Endo et al., 2013乯丅偙傟傜偺寢壥偼丄偙傟傑偱晄柧偱偁偭偨僼傿僩僋儘儉偵傛傞壴夎宍惉惂屼偺暘巕婡峔偵娭偡傞廳梫側抦尒偱偁傞丅
丂丂 僼傿僩僋儘儉 尋媶top傊
|
丂丂偙傟傑偱偵峴偭偰偒偨僼傿僩僋儘儉暘巕偺峔憿乛婡擻夝愅傗丄嵟嬤偺僼傿僩僋儘儉乮僶僋僥儕傾丒僼傿僩僋儘儉乯偺棫懱峔憿偺夝柧偵傛傝丄僼傿僩僋儘儉暘巕忋偺偳偺傛偆側峔憿偑偳偺傛偆側婡擻偲娭傢傞偐傪帵偡峔憿乛婡擻儅僢僾偑柧傜偐偵側傝偮偮偁傞丅偙傟傪棙梡偟偰丄嫗搒戝妛丒悾屗岥峗彶攷巑偺僌儖乕僾丄壀嶳戝丒抮揷孾攷巑傜偲嫟摨偱丄僼傿僩僋儘儉偺帺慠曄堎偵婡擻揑側堄媊晅偗傪偡傞偙偲傪帋傒偰偄傞丅
丂丂 僼傿僩僋儘儉 尋媶top傊
|
|